|
Научная работа: Выделение белковНаучная работа: Выделение белковВЫДЕЛЕНИЕ БЕЛКОВ Выделение практически чистого индивидуального белка (в таких случаях нередко употребляют не вполне удачный термин "гомогенный белок") — необходимая предпосылка для изучения его строения и функциональных свойств. Технические приемы, используемые для этого, весьма многообразны и быстро совершенствуются, причем наряду с развитием микрометодов все чаще возникает необходимость масштабирования процессов, с тем, чтобы получать крупные количества высокоочищенных белков для нужд медицины и биотехнологии. Особенно обострилась потребность в эффективных методах разделения белков с развитием генно-инженерных способов их получения. В данной главе рассмотрены лишь наиболее общие принципы препаративной химии белков.
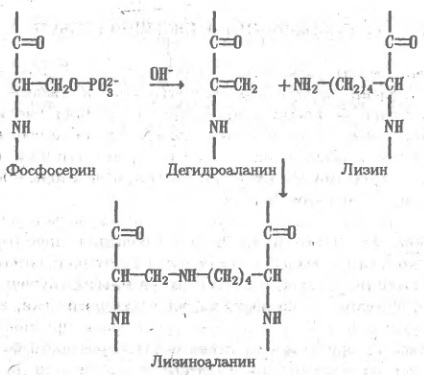
Особенности выделения белков Получение чистых химических индивидуальных белков включает в себя как удаление небелковых примесей, так и разделение между собой собственно белковых компонентов. Последняя часть задачи в силу сходства физико-химических свойств белков особенно сложна, поэтому именно ее решение определяет выбор тех или иных схем выделения белка. При этом необходимо учитывать некоторые особенности поведения, присущие всем белкам. К ним относится, прежде всего, способность белков подвергаться денатурации, т.е. претерпевать такие изменения пространственного строения, которые приводят к утрате или частичной потере функциональных свойств. Правда, денатурация во многих случаях обратима, однако эта обратимость не обеспечивается автоматически, а требует в каждом отдельном случае подбора специальных приемов. В то же время полностью или даже частично денатурированные белки весьма уязвимы для необратимых повреждений, в особенности для действия протеолитических ферментов, поэтому условий, способствующих денатурации, следует всемерно избегать. Для предотвращения тепловой денатурации выделение белка проводят при низкой температуре, обычно при 4 ºС. Необходимо также избегать крайних значений pH. Белки легко денатурируют при низких рН из-за протонирования отрицательно заряженных в нормальных условиях карбоксильных групп и возникающего вследствие этого резкого преобладания положительных зарядов, которое благоприятствует развертыванию компактной структуры. В щелочной среде при рH 10 и выше утрачиваются положительные заряды, обусловленные протонированием ε-аминогрупп лизина, и опять-таки наступает декомпенсация зарядов на поверхности, дестабилизирующая глобулу. Одновременно фенольные гидроксилы тех остатков тирозина, которые были скрыты внутри глобулы, получив отрицательный заряд, стремятся выйти на поверхность, что также способствует денатурации белка. В еще более щелочных растворах происходят реакции, необратимо повреждающие белок, — отщепление серы от остатков цистеина и цистина, β-элиминирование фосфатных остатков в фосфопротеинах, отщепление полисахаридных цепей, присоединенных к остаткам серина в гликопротеинах. Образующиеся в результате этих реакций остатки непредельной аминокислоты — дегидроаланина — присоединяются к близлежащим ε-аминогруппам лизина с образованием так называемого лизиноаланина — соединения, присутствие которого в кислотном гид-ролизате белка свидетельствует о глубоком повреждении его структуры.
Причиной денатурации может стать применение органических растворителей, способных нарушить существенную для стабильности белка систему гидрофобных контактов внутри глобулы. Надо также учитывать возможность денатурации белка на границе раздела фаз, В особенности при образовании пены. При выделении очень малых количеств белка, не редком в современной биохимии, следует считаться с потерями, вызываемыми его адсорбцией, часто необратимой, на поверхности стекла или полимерных материалов, особенно значительной на заключительных стадиях очистки. Особую опасность при выделении белков представляют содержащиеся в исходном материале протеолитические ферменты, даже если они присутствуют в следовых количествах. Повреждение, которое приводит к образованию ложных множественных форм белка, а иногда и его глубокое расщепление становятся особенно вероятными, если схема выделения включает условия, приближающие белок к денатурации. Для предотвращения протеолиза необходимо уже на ранних стадиях очистки включать операции, направленные на отделение протеиназ, прибавлять к смеси ингибиторы различных классов протеиназ. Очень важно проводить очистку белка быстро, что снижает опасность протеолиза и вероятность денатурации. Распространено мнение об особой нестабильности чистых белков. Действительно, содержащиеся в неочищенных препаратах посторонние белки в определенной мере защищают белок от денатурирующих воздействий и протеолиза. На промежуточных стадиях, когда полная очистка еще не достигнута, опасность протеолиза увеличивается, с удалением же следов протеиназ на завершающих этапах очистки стабильность белка должна вновь возрасти. Впрочем, возрастает и опасность сорбционных потерь на поверхностях, которые могут восприниматься экспериментатором как присущая чистому белку нестабильность. Характерные свойства белков, на которых основано их разделение
На каких физико-химических особенностях белков могут быть основаны способы их разделения? Во-первых, это размер молекулы, ее геометрия. На использовании этой особенности базируются методы гель-хроматографии и ультрафильтрации, отчасти электрофорез в гелях. Во-вторых, характерное для данного белка распределение заряженных групп на его поверхности. Соотношение катионных и анионных групп в белке меняется в зависимости от рН, изоэлектрические точки белков — pI (значения рН, при котором положительные и отрицательные заряды белка полностью компенсированы и суммарный заряд равен нулю) существенно различаются у разных белков. Известны белки, являющиеся в физиологических условиях катионными, анионными или молекулами без заметного преобладания того или иного заряда. На различии заряда белков при разных рН основано их разделение методами электрофореза, изоэлектрического фокусирования, изоэлектрической и ионообменной хроматографии. Существенно, однако, не только соотношение заряженных групп, определяющее значение pI. Белки со сходными изоэлектрическими точками могут различаться распределением заряженных функциональных групп по поверхности глобулы. Последние размещаются более или менее равномерно либо; наоборот, образуют локальные сгущения, гроздья одинаково заряженных групп, что сказывается при ионнообменной хроматографии белка. В-третьих, белки различаются числом и характером гидрофобных участков поверхности, что используют при гидрофобной хроматографии Заметим, что ни один из рассмотренных выше признаков не может сам по себе обеспечить выделение индивидуального белка из сложной смеси — они недостаточно характеристичны, не гарантируют избирательности очистки. Значительно более перспективно в этом отношении использование для выделения функциональных свойств белка. Действительно, среди множества белков в исходном материале найдется немало таких, которые имеют сходную молекулярную массу или близкие изоэлектрические точки, однако число, например, фосфатаз или амилаз будет заведомо небольшим. Очевидно, что метод выделения, основанный на использовании способности этих ферментов взаимодействовать со своим специфическим субстратом, несравненно избирательнее, чем любой прием, базирующийся на разнице физико-химических свойств. Схемы выделения белков, использующие только один какой-либо принцип, редки, обычно различные подходы к фракционированию сочетаются и дополняют друг друга. Разделение белков по молекулярной массе. Гель-хроматография (гель-фильтрация)
В этом методе используют гранулированные гели поперечно-сшитых гидрофильных материалов, например декстрана (сефадекс, сефароза и их аналоги), полиакриламида (биогели и их аналоги), поливинилового спирта (тойоперл). Гранулы образованы трехмерной сеткой полимера, которая непроницаема для крупных молекул, частично проницаема для молекул промежуточного размера и хорошо проницаема для небольших молекул, солей и воды. В зависимости от среднего размера ячейки полимерного геля и геометрии молекулы последней доступна большая или меньшая часть общего объема гранул геля. При движении раствора, содержащего белки и другие молекулы, по
колонке, которая заполнена набухшими гранулами геля, компоненты смеси,
проникшие в гель, задерживаются в нем. Таким образом, они отстают от более
крупных молекул, которые не могут войти внутрь гранул и находятся только в
омывающем их растворе. Не будучи включенными в гель, крупные молекулы появляются
в элюате, как только через колонку пройдет "свободный" объем раствора
Если размеры молекул белка таковы, что они могут проникать в поры, составляющие некоторую часть объема гранул, то будет наблюдаться задержка элюции и белок появится в объеме Ve, связанном с коэффициентом доступности (долей объема гранул, доступной данному виду молекул) соотношением: где V, — полный объем колонки, за вычетом той его части, которая приходится на сам гель образующий полимер. Каждому белку в зависимости от размеров его молекулы соответствует
свое значение, на чем и основало разделение при гель-хроматографии. Понятно,
что если объем элюции Строго говоря, при гель-хроматографии разделение белков определяется не молекулярной массой, а геометрическими размерами молекулы. Соответственно, молекулы вытянутой формы за счет "кувыркания" в растворе труднее проникают в гели, чем сферические молекулы такой же молекулярной массы. Этим объясняется ранняя элюция денатурированных белков, которые ведут себя как неупорядоченный рыхлый клубок, а не как компактная глобула. Простая зависимость между объемом элюции и молекулярной массой белка (справедливая, конечно, только для компактных сферических молекул) и легкость эксперимента сделали гель-хроматографию излюбленным методом определения молекулярной массы белков. Для этой цели колонку, заполненную соответствующим гелем, калибруют набором белков с известными молекулярными массами, после чего определяют объем элюции изучаемого белка и вычисляют его молекулярную массу интерполяцией. Точность метода не очень велика, но вполне достаточна для большинства практически встречающихся задач.
При использовании данного метода необходимо учитывать ограничения, возникающие из-за того, что гель, образующий материал не вполне инертен, как это предполагает теория метода, и может взаимодействовать с разделяемыми веществами, что искажает зависимость объема элюции от размера молекулы. Это особенно сказывается при разделении малых количеств белка, так как сорбционная емкость матрицы геля невелика и в крупномасштабных опытах ее взаимодействие с белком мало отражается на процессе. Связывание белков гельобразующими материалами может быть вызвано ионообменными взаимодействиями, в частности содержанием отрицательно заряженных групп в полисахаридных матрицах (сефароза, сефадекс), а также в полиакриламидных материалах. В последних карбоксильные группы возникают при спонтанном гидролизе амидов, в полисахаридах же они могут образовываться в результате окисления. Задержка при гель-хроматографии, вызванная ионообменными взаимодействиями с матрицей, особенно характерна для катионных белков, например лизоцима и некоторых субтилизинов. Нередко она весьма значительна и может даже препятствовать отделению солей от белка. В аналитических опытах такое удерживание удается подавить значительным повышением ионной силы раствора. Еще одна причина аномального удерживания веществ при гель-хроматографии, особенно заметная при выделении небольших молекул, например пептидов. — гидрофобное связывание с матрицей геля. Гидрофобные элементы включаются в гидрофильные полисахаридные матрицы при обработке их сшивающими агентами, в частности эпи-хлоргидрином, при синтезе сефадекса. Пептиды, содержащие гидрофобные, в особенности ароматические, остатки (фенилалаиина, триптофана) иногда удерживаются матрицей столь значительно, что появляются в элюате позже неорганических солей. Разрешающая способность метода не очень велика, в то же время простота проведения и мягкость условий эксперимента являются его бесспорными преимуществами. Применимость метода на первых этапах очистки ограничивается тем, что для удовлетворительного фракционирования белков объем наносимого раствора не должен превышать 3-5% общего объема колонки. Ввиду этого к гель-хроматографии обычно прибегают в середине или на завершающих этапах выделения белка. Разумеется, при отделении низкомолекулярных примесей, в частности при обессоливании, объем образца может быть значительно большим, поскольку не требуется высокого разрешения. В таком упрощенном варианте гель-фильтрацию используют особенно часто. Несмотря на указанные ограничения, гель-хроматография — удобный способ фракционирования белков. Его применяют и для отделения от белков низкомолекулярных примесей, в том числе солей. В последнее время наряду с гельобразующими материалами для разделения белков по размерам начали применять макропористые неорганические носители макропористые стекло и силикагель. Обычно поверхность этих материалов покрывают гидрофильными органическими веществами, чтобы исключить необратимую сорбцию белков. Жесткость этих материалов позволяет разделять белки по размеру молекул при повышенных давлениях, что ускоряет процесс и снижает помехи со стороны диффузии. Ионообменная хроматография белков
Метод ионообменной хроматографии, основанный на различиях в соотношении и распределении заряженных групп на поверхности белка, принадлежит к числу наиболее используемых. В хроматографии белков практически не применяют синтетические ионообменные смолы на основе полистирола, весьма популярные в аналитической химии аминокислот и пептидов. Это объясняется, во-первых, большим содержанием поперечных сшивок, делающих материалы такого рода практически непроницаемыми для белков, во-вторых, сорбцией белков, подчас необратимой, на гидрофобной поверхности полистирола. По указанным соображениям для разделения белков используют ионообменники, в которых матрица ("подложка") отчетливо гидрофильна. Особенно распространены ионообменники, получаемые присоединением монотонных групп к целлюлозе, поперечно-сшитым декстранам (сефадексы). Для получения катионов в качестве ионогенной чаще всего используют карбоксильную группу рКа которой, несколько изменяющийся в зависимости от микроокружения, близок к 4 (карбоксиметил (СМ)-производные целлюлозы, сефадекса, содержащие группировки
Карбоксильные группы таких ионитов отрицательно заряжены при рН 5 и выше и, следовательно, способны связывать белки, которые в этих условиях несут положительный заряд. Связывание белков усиливается, если на их поверхности встречаются скопления ("гроздья") катионных групп. При прочих равных условиях с катионитом лучше связываются белки большей молекулярной массы, что объясняется кооперативностью многоточечного взаимодействия обширных участков поверхности такого белка с анионными группами ионообменника. Десорбция белков, связанных катионитом, обычно достигается повышением ионной силы элюирующего раствора, причем взаимодействующие между собой заряженные группы белка и ионита оказываются в окружении противоположно заряженных ионов солей. В результате при определенной концентрации соли, характерной для каждого белка, электростатические взаимодействия между ним и ионитом снимаются и белок элюируется с колонки. Плавное увеличение ионной силы раствора, применение линейного или более сложного градиента концентрации соли вызывает десорбцию сначала наиболее слабо удерживаемых молекул, затем более прочно связанных белков и т.д. В препаративных опытах нередко прибегают к ступенчатой элюции, при которой концентрации соли повышается скачками. Это ускоряет разделение и позволяет собрать белок в небольшом объеме, однако легко приводит к образованию одним и тем же белком нескольких ложных. При промывании колонки с ионитом раствором соли подходящей концентрации белок десорбируется, иногда образуя довольно длинный "хвост", что может быть следствием неравномерного распределения ионных групп в сорбенте. Участки с их повышенным содержанием, скопления таких групп прочнее удерживают белок, что и вызывает задержку элюции и образование "хвоста". В такой ситуации скачкообразное повышение ионной силы элюирующего раствора резко улучшает условия десорбции, поэтому часть белка, которая в обычных условиях образовывала бы "хвост", десорбируется скачком, давая ложный пик. Ввиду этого следует определять белковый состав каждой фракции независимым методом, например электрофорезом в полиакриламидном геле, или подвергать сомнительные пики повторной хроматографии в тех же условиях. Несоблюдение таких предосторожностей нередко приводит к ошибочному обнаружению "множественных форм" белков. В принципе для десорбции белков с катионитов можно прибегать и к градиенту рН. Например, понижение pH до 3—4 приводит к протонированию карбоксилатных ионов карбоксиметилцеллюлозы или аналогичных ионитов и постепенной десорбции связанных с ними белков. Можно рассчитывать и на достижение изоэлектрической точки сорбированного белка, что опять-таки вызвало бы его десорбцию. Однако такой прием используют не часто не только из-за опасности денатурации белков при понижении рН, но и из-за осложнений, связанных с сорбцией ионитом части ионов элюирующего раствора и неопределенностью вызываемых этим локальных изменений рН. По таким же соображениям не рекомендуется использовать в ионообменной хроматографии белков растворы, содержащие несколько разных катионов или анионов. Помимо карбоксильных катионитов, о которых шла речь выше,
применяют катиониты, содержащие сульфогруппы — сульфопропилсефадекс. В отличие от карбоксильных сульфогруппы сохраняют отрицательный заряд практически при всех рН, используемых в хроматографии белков. Ионогенные группы фосфоцеллюлозы слабее, чем у сульфопропилсефадекса, но сильнее карбоксильных. Оказалось, что этот ионообменник может применяться в качестве биоспецифического сорбента при выделении некоторых ферментов фосфорного обмена. Среди анионитов наибольшее распространение получили диэтил-амииоэтил (DEAE)-целлюлоза и ее аналоги с иными матрицами: Диэтиламиноэтильная группа, присоединенная к целлюлозе или другой полисахаридной матрице. Основность диэтиламиноэтильной группы в сорбентах этого типа
несколько ниже обычной из-за влияния окружающих ее гидроксильных групп, так что
ее р Хроматография белков иа DAEA-целлюлозе и аналогичных аниони-тах, подобно хроматографии на катионах, определяется многоточечным связыванием отрицательно заряженных групп белка (прежде всего карбоксильных) с катионными группами ионообменника. Как и при хроматографии на катионитах, десорбции белков достигают повышением ионной силы элюирующего раствора, ее можно проводить ступенчато или с применением градиента концентрации соли. И в этом случае не рекомендуют градиенты pH и использование растворов, содержащих разные анионы. Например, при десорбции белков хлористым натрием с DEAE-целлюлозы, уравновешенной ацетатным буфером, помимо экранирования разноименно заряженных групп сорбента и белка происходит замещение удерживаемых анионитом ацетат-ионов ионами хлора. Если элюция проводится в слабокислых растворах, десорбированные ацетат-ионы, связывая протоны, вызовут сдвиг рH в щелочную сторону, что приведет к локальному изменению условий элюции и сделает процесс трудно контролируемым. В последнее время в хроматографии белков все шире применяют ионообменники на основе гидрофильных органических полимеров, получаемых в форме шариков строго одинакового диаметра (10 мкм) и обладающих большой рабочей поверхностью. Стандартность гранул таких ионитов снижает размывание хроматографических пиков за счет различий во времени диффузии белковых молекул внутри сорбента — фактор, ограничивающий эффективность обычных ионообменников с неодинаковыми гранулами. Носители этого типа жестки, что позволяет достигать весьма высоких скоростей протекания растворов через колонку при давлении порядка 20 атм. Совместное действие этих факторов резко повышает эффективность и скорость хроматографии белков на такого рода сорбентах, получившей название быстрой жидкостной хроматографии белков (английское сокращенное обозначение FPLC). В качестве сорбентов используют Moho-Q — сильный анионит с четвертичными аммонийными группами, Moho-S — сильный катионит, содержащий сульфогруппы, или Моно-P— катионит с фосфатными группами.
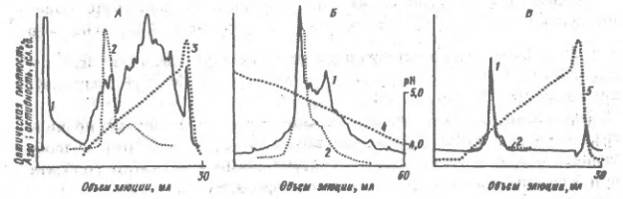
Очистка щелочной фосфатазы ионообменной хроматографией: 1 - оптическая плотность при 280 ни, отражающая общее содержание белка; 2 - активность щелочной фосфазы; А - разделения неочищенного препарата на аиионите Моно-Q при рН 8 в градиенте концентрации NaCl 0-0.35 М (3): Б - хроматофокусирование на Моно-Р щелочной фосфатазы, очищенной на предыдущей стадии в градиенте рН (4); В - хроматография фракции, выделенной на Моно-Р, на анионите Моно-Q в градиенте концентрации NaCl (5) К сожалению, до сих пор не удалось разработать достаточно эффективный способ препаративного электрофореза белков в гелях, хотя их аналитическое разделение электрофорезом в полиакркламидном геле дает очень хорошие результаты и является ведущим методом в исследовании белковых смесей и индивидуальных белков. Гидрофобная хроматография белков
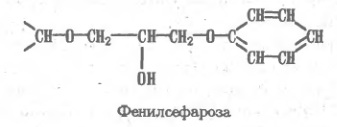
Как известно, поверхность белковой глобулы богата гидрофильными аминокислотами, но в то же время содержит немало (до половины от их общего содержания) гидрофобных остатков, нередко образующих скопления ("гроздья"). Такие гидрофобные зоны, развитые в большей или меньшей степени, представляют характерную особенность структуры каждого белка, на чем и основан метод гидрофобной хроматографии. Соответствующие сорбенты синтезируют, включая гидрофобные группировки в гидрофильную матрицу, например в поперечно-сшитую агарозу — сефарозу. По такому принципу построены, в частности, октил- и фенилсефароза:
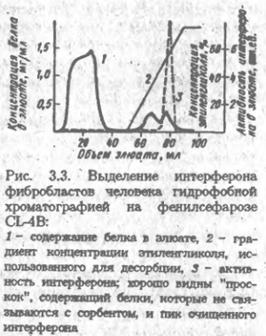
При пропускании белкового раствора через фенилсефарозу гидрофобные участки поверхности белков образуют контакты с фенильными группами, вытесняя прилегающие к этим структурам молекулы воды. Число и прочность таких контактов весьма различны у разных белков. Повышению их прочности способствует сорбция белков из концентрированных растворов солей, например сульфата аммония. Плавное понижение концентрации соли в растворе, протекающем через колонку с фенилсефарозой, приводит к поочерёдной десорбции белков. Сходным образом действуют сорбенты, полученные присоединением к макропористому кремнезему гидрофобных алкильных радикалов различной длины. Они, будучи жесткими, особенно подходят для
работы при повышенном давлении в условиях высокоэффективной жидкостной
хроматографии (ВЭЖХ, англ. HPLC). Те из них, которые содержат длинные — Нередко гидрофобная хроматография сочетается с другими эффектами. Например, присоединение к активированной бромцианом сефарозе диаминов различной длины дает сорбенты, в которых содержатся гидрофобные углеводородные цепочки наряду с двумя катионными группами. Объединение черт гидрофобного сорбента и анионита в одном хроматографическом материале обогащает его. Описанный выше способ проведения хроматографии на гидрофобном сорбенте — далеко не единственно возможный. Для сорбции белков не обязательно введение в раствор повышенных концентрация соли, а для элюцни можно применять добавление органических растворителей, сдвиг рН. В некоторых случаях, когда связывание белка основано на сочетании гидрофобных и ионных взаимодействий, хорошие результаты дает элюция растворами солей. Отметим также, что признаки гидрофобной хроматографии встречаются и в других приемах разделения белков, в особенности в аффинной хроматографии. Аффинная, или биоспецифическая. Хроматография белков
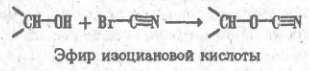
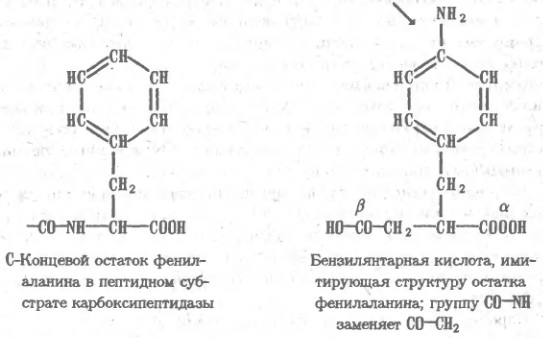
Как уже отмечалось, способы разделения белков, основанные на различиях в их физико-химических свойствах, не могут быть высокоспецифичными. С этой точки зрения гораздо перспективнее препаративные методы, которые базируются на функциональных различиях белков. Для множества белков, включая ферменты, их ингибиторы, транспортные белки, иммуноглобулины, регуляторные белки, токсины, рецепторы, — главнейшая функциональная черта состоит в способности избирательно связывать те или иные лиганды субстраты, коферменты, аллостерические эффекторы, антигены и т.п. Такое связывание по существу своему весьма специфично, и это свойство резко выделяет тот или иной белок из множества других. На использовании функционально обусловленной способности белков обратимо связываться с соответствующими лигандами и основан метод аффинной, или биоспецифичгсгой. хроматографии. Для синтеза аффинного сорбента отвечающий специфичности данного белка лиганд (субстрат или его аналог при хроматографии фермента) присоединяют к инертной матрице так, чтобы возможно меньше затронуть те элементы его структуры, которые непосредственно участвуют во взаимодействии с белком. Иногда лиганд прямо вводят в реакцию с функциональными группами на поверхности матрицы, однако чаше их соединяют через промежуточное звено ("вставку", "ножку" и т.п.), чтобы отдалить лиганд от матрицы и уменьшить пространственные препятствия для его сближения с белком. К каждому из элементов структуры аффинного, сорбента предъявляются определенные требования. Матрица должна быть инертной и не создавать стерических препятствия для белка. Чаще всего в качестве матрицы используют микропористые гели, образованные поперечно-сшитыми гидрофильными полимерами, например производное агарозы — сефарозу, синтетические полимеры или неорганические носители макропористые производные кремнезема — силикагель или стекло. Присоединение лигандов к сефарозе (непосредственно или через вставку) обычно проводят, активируя ее бромцианом. Бромциан (сильный яд!) в щелочкой среде реагирует с гидроксильными группами сефарозы, образуя весьма активный эфир изоциановой кислоты: Последний взаимодействует затем с аминогруппами лиганда L или "вставки" с образованием производного изоыочевнны, которое является сильным основанием и в обычном диапазоне рН песет положительный заряд: Таким образом, присоединение лиганда к сефарозе, активированной бромцианом, одновременно создает на матрице эквивалентное содержанию лиганда число катионных групп. Следовательно, такой сорбент помимо био- специфического связывания белка лигандом может проявлять свойства анионита, что необходимо учитывать при его использовании. Известны и другие способы присоединения лнгандов к матрице, однако в каждом случае следует учитывать изменения в характере носителя, сопровождающие реакцию с лигандом, а также большую или меньшую неустойчивость образовавшейся связи, которая иногда может вызывать постепенную "утечку" лиганда. Например, изомочевина может превращаться в производное гуанидина в присутствии значительных концентраций первичных аминов или аммиака, что влечет за собой отщепление лиганда. Сложнее требования, предъявляемые к лиганду. Он прежде всего должен достаточно сильно взаимодействовать с белком. Так, принято считать, что для получения сорбентов, предназначенных для выделения ферментов, в качестве лигандов могут быть использованы такие ингибиторы или аналоги субстрата, которые имеют константу ингибировавия (субстратную константу), т.е. константу диссоциации фермент — лиганд, не хуже 10-4 М. При этом учитывается, что присоединение лиганда к матрице ухудшает (т.е. повышает) константу ингибирования по крайней мере на один порядок. При подборе лигандов для аффинной хроматографии ферментов приходится изменять структуру субстрата, чтобы предотвратить возможность его превращения ферментом в продукт, например, расщепления пептидного лиганда, если речь идет о хроматографии протеиназ. Очевидно, что сорбент, содержащий в качестве лиганда истинный субстрат, будет трансформирован при первом же контакте с ферментом и окажется непригодным. При пропускании раствора, содержащего сложную смесь белков, других биополимеров, низкомолекулярных соединений, солей, пигментов и т.п., через биоспецифический сорбент, построенный по описанной выше схеме, лнганд образует комплекс только с тем белком, который имеет участок связывания, комплементарный структуре лиганда. В результате именно этот белок удерживается аффинной колонкой, тогда как все остальные компоненты смеси проходят через нее не задерживаясь. После промывания колонки для удаления неспецифически удерживаемых примесей белок элюируют, изменяя состав протекающего через колонку раствора так, чтобы ослабить взаимодействие белка с лигандом. Для этого прибегают к изменению рН, добавляют к элюенту неорганические соли, органические растворители. Все эти факторы, воздействуя на структуру зоны связывания лигвнда и подавляя от дельные виды белок-лиг видных взаимодействий, например ионные связи и гидрофобные контакты, обеспечивают десорбцию. В отдельных случаях прибегают к аффинной элюции раствором лиганда или его аналога. Этот прием, основанный ив конкуренции за связывание белка между присоединенным к матрице и растворенным лигандами, весьма специфичен и эффективен, но дорог. В качестве примера рассмотрим применение циклопептидного антибиотика — грамицидина S — в качестве лиганда при аффинной хроматографии протеолитических ферментов. Он содержит ряд гидрофобных аминокислот, соответствующих специфичности многих протеиназ, и два остатка орнитина, δ-аминогруппы которых позволяют легко присоединить циклопептид к активированной бромцианом сефарозе или другим матрицам: Остаток орнитина, аминогруппа, которого присоединена к активированной бромцианом сефарозе. Грамицидин S связывает протеиназы различных классов, однако он устойчив к их действию и не подвергается гидролизу, по-видимому, из-за присутствия в структуре остатков D-фенилаланина, пролина, а также из-зa особенностей конформации циклопептида. Следовательно, грамицидин S является природным аналогом субстратов протеназ и хорошо подходит для роли лигаида широкой специфичности. Так, при пропускании через колонку с грамицидин-5-сефарозой сложной смеси веществ, содержащихся в культуральной жидкости одного из штаммов бактерии Bacillus subtilis, сорбент связывает только металлопротеиназу, которую удается полностью освободить от примесей и получить практически чистый белок. Еще эффективнее сорбенты, содержащие такие лиганды, которые взаимодействуют не только с зоной связывания, но и с каталитическим центром фермента. Например, производные бензилянтариой кислоты оказались хорошими лигандами для выделения карбоксипептидаз — ферментов, отщепляющих аминокислоты от карбоксильного конца пептидной цепи. Сходство лигандов с субстратами карбоксипептидаз на первый взгляд невелико, однако оказывается, что группа HООC-
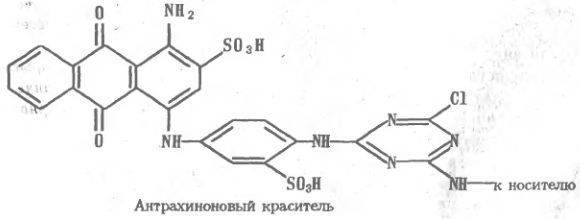
В результате Широко используют своеобразный вариант аффинной хроматографии, основанный на применении в качестве лигандов синтетических красителей антрахинонового ряда, например цибакрона голубого;
Плоская структура, образованная тремя конденсированный и ароматическими кольцами замещенного антрахинона, к которому в положении 1 присоединены замещенный анилин и триазиновый цикл, способна избирательно взаимодействовать с целым рядом белков. Лиганд связывается, по-видимому, во впадинах, которые нередки на поверхности белков, в особенности в активных центрах ферментов. Высказывались предположения, что антрахиноны вместе с присоединенными ядрами анилина и триазина образуют структуру, стерически близкую природным лигандам, например таким коферментам, как NAD, и имитируют свойственный последним способ связывания с белками. Ввиду этого сорбенты, содержащие цибакрон голубой и некоторые другие антрахиноновые красители, особенно рекомендуются для аффинной хроматографии белков, содержащих NAD или иные соединения нуклеотидной природы. Нет необходимости, чтобы нуклеотид или сходное по строению соединение было похоже на субстрат — оно может имитировать эффектор, регулирующий активность фермента. Так, например, хорошие результаты дает хроматография на антрахиноновых красителях аспартаткарбомилазы, где нуклеотиды выступают как аллостерические эффекторы. Впрочем, круг белков, связываемых этими сорбентами, значительно шире и включает, например, сывороточный альбумин. Последний легко присоединяет различные лиганды, в том числе некоторые лекарственные препараты и триптофан, который можно (с большими оговорками) считать напоминающим конденсированные кольца антрахиноиа. Структура лиганда— замещенного антрахинона — не безразлична, поэтому, используя разные красители, удается получить серию сорбентов, не одинаково связывающих различные белки. Иммуносорбция
К числу наиболее специфических принадлежит взаимодействие белков с антителами — иммуноглобулинами. Это свойство с успехом используют для создания биоспецифических сорбентов, в которых роль лиганда выполняют специфические в отношении выделяемого белка иммуноглобулины. Получение и применение такого рода сорбентов происходят примерно так же, как и в обычной аффинной хроматографии, отличаясь лишь немногими особенностями. Во-первых, присоединение иммуноглобулина G к матрице необходимо проводить в щадящих условиях. Установление между молекулой антитела и носителем множества ковалентных связей (которое может произойти, если добиваться повышения емкости сорбента) благоприятствует денатурации белка и может, следовательно, привести к утрате специфичности антитела. Во-вторых, использование обычных, поликлональных, антител, которые представляют собой весьма сложную смесь молекул иммуноглобулинов различного строения, приводит к получению сорбента, в котором содержатся антитела с широким диапазоном сродства к выделяемому белку-антигену. Это затрудняет его сорбцию и в особенности десорбцию, так как диссоциация целого набора неодинаковых комплексов антиген—антитело может потребовать вариации условий алюции в весьма широком интервале, вплоть до весьма жестких. Это осложнение снимается, если доступны моноклональные антитела, представляющие собой однородные молекулы иммуноглобулина. Десорбция и в этом случае может доставлять некоторые трудности из-за прочности комплекса, однако их удается преодолеть, подавляя нековалентные взаимодействия между выделяемым белком и иммуноглобулином при помощи изменения рН, ионной силы, добавления в элюент органических растворителей или мочевины. Иммуносорбция особенно уместна как высокоспецифичный метод выделения небольших количеств особо ценных белков из сложных смесей, поскольку используемые сорбенты дороги и недолговечны из-за склонности иммуноглобулинов к денатурации. Очень важно, что к иммуносорбции можно прибегнуть даже в таких случаях, когда сведения о свойствах белка, в том числе и о его функции, скудны. Если известна первичная структура, определенная по нуклеотидной последовательности гена, то возможно синтезировать ряд фрагментов его пептидной цепи и, присоединив их к белку-носителю, иммунизировать таким конъюгатом животное. Это дает возможность получить антитела к исследуемому белку, несмотря на то что он не только не выделен, но и его функция пока неизвестна. Иммуносорбенты, синтезируемые присоединением таких антител к подходящему носителю, позволяют выделить белок. Перспективы использования белковой инженерии для выделения белков Методы генной инженерии позволяют присоединять к структурному гену белка последовательности, кодирующие такие белки или их фрагменты (домены), которые облегчают выделение конструкции в целом. Полученный гибридный ("химерный") белок во многих случаях, хотя и не всегда, жизнеспособен, причем обе части "химеры" не зависимо друг от друга свертываются в компактные структуры. Это дает возможность, используя характерные свойства присоединенной к целевому белку структуры, провести специфическое выделение гибрида, после чего расщепить "шарнир", соединяющий обе его части. Так, белки-гибриды, в которых одной из составных частей является белок А стафилококков, могут быть очищены хроматографией на колонках, содержащих присоединенные ковалентно иммуноглобулины. Белок А прочно связывается с последними. Гибридные белки, в состав которых входит β-галактозидаза или ее крупные фрагменты, могут быть очищены на колонках, специфически сорбирующих этот фермент, например содержащих ковалевтно связанный аналог субстрата — амннофенилтиогалактозид. По завершении очистки гибридный белок подвергают ограниченному протеолизу или расщепляют связь между объединенными в его структуре белками специфическими химическими методами, например действием бромциана, и отделяют "целевой" белок от белка-носителя, что, как правило, не составляет затруднений. Иногда к структурному гену белка присоединяют олигонуклеотидные фрагменты, которые кодируют короткие аминокислотные последовательности, наращивающие этот белок с аминного или карбоксильного конца. Эти последовательности выбирают так, чтобы они не препятствовали свертыванию белка и в то же время сообщали всей конструкции то или иное характерное свойство, которое позволяет легко выделить такую молекулу даже из очень сложной смеси. Затем эти фрагменты отщепляют в мягких условиях. Так, присоединение к карбоксильному концу дегидрофолатредуктазы Lys—Gly—Ser—Arg—Ser двух остатков гистидина дает последовательность Lys—Gly—Ser—Arg Ser—His—His. Соседствующие остатки гистидина образуют комплекс с ионом Ni2+, который удерживается специально полученным сорбентом с хелатирующими группами. Это позволяет в одну стадию выделить клонированный E.coli фермент из культуральной жидкости практически чистым, с 90%-ным выходом. Отщепление обоих остатков гистидина карбоксипептидазой А приводит к дегидрофолатредуктазе с G-концевой последовательностью Lys-Gly-Ser-Arg. Предложено к аминному концу рекомбинантных белков-лимфокинов присоединять последовательность Asp—Туr—Lys—Asp—Asp—Asp— Аsр—Аsр—lys, которую связывает специально полученное антитело. После очистки белка (модифицированного присоединением такой последовательности) имнуносорбцией его обрабатывают энтеропептидазой — ферментом, специфически отщепляющим этот пептид. По-видимому, описанный подход особенно перспективен для выделения и очистки рекомбинантных белков. |
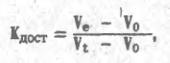

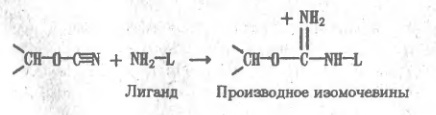
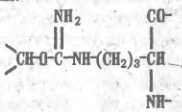

|
|
| 17.06.2012 |
| Большое обновление Большой Научной Библиотеки |
| 12.06.2012 |
| Конкурс в самом разгаре не пропустите Новости |
| 08.06.2012 |
| Мы проводим опрос, а также небольшой конкурс |
| 05.06.2012 |
| Сена дизайна и структуры сайта научной библиотеки |
| 04.06.2012 |
| Переезд на новый хостинг |
| 30.05.2012 |
| Работа над улучшением структуры сайта научной библиотеки |
| 27.05.2012 |
| Работа над новым дизайном сайта библиотеки |